Нервная ткань. Периферическая нервная система (развитие, состав) Основные вопросы темы
ОСНОВНЫЕ ВОПРОСЫ ТЕМЫ:
1. Общая морфофункциональная характеристика нервной ткани.
2. Эмбриональный гистогенез. Дифференцировка нейробластов и глиобластов. Понятие о регенерации структурных компонентов нервной ткани.
3. Нейроциты (нейроны): источники развития, классификация, строение, регенерация.
4. Нейроглия. Общая характеристика. Источники развития глиоцитов. Классификация. Макроглия (олигодендроглия, астроглия и эпендимная глия). Микроглия.
5. Нервные волокна: общая характеристика, классификация, строение и функции безмиелиновых и миелиновых нервных волокон, дегенерация и регенерация нервных волокон.
6. Синапсы: классификации, строение химического синапса, строение и механизмы передачи возбуждения.
7. Рефлекторные дуги, их чувствительные, двигательные и ассоциативные звенья.
ОСНОВНЫЕ ТЕОРЕТИЧЕСКИЕ ПОЛОЖЕНИЯ
НЕРВНАЯ ТКАНЬ
Нервная ткань выполняет функции восприятия, проведения и передачи возбуждения, полученного из внешней среды и внутренних органов, а также анализ, сохранение полученной информации, интеграцию органов и систем, взаимодействие организма с внешней средой.
Основные структурные элементы нервной ткани – клетки и нейроглия .
Нейроны
Нейроны состоят из тела (перикариона ) и отростков, среди которых выделяют дендриты и аксон (нейрит). Дендритов может быть множество, аксон всегда один.
Нейрон как любая клетка состоит из 3 компонентов: ядра, цитоплазмы и цитолеммы. Основной объём клетки приходится на отростки.
 Ядро
занимает центральное положение в перикарионе.
В ядре хорошо развито одно или несколько ядрышек.
Ядро
занимает центральное положение в перикарионе.
В ядре хорошо развито одно или несколько ядрышек.
Плазмолемма принимает участие в рецепции, генерации и проведении нервного импульса.
Цитоплазма нейрона имеет различное строение в перикарионе и в отростках.
В цитоплазме перикариона находятся хорошо развитые органеллы: ЭПС, комплекс Гольджи, митохондрии, лизосомы. Специфичными для нейрона структурами цитоплазмы на светооптическом уровне являются хроматофильное вещество цитоплазмы и нейрофибриллы .
 Хроматофильное вещество
цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при окрашивании нервных клеток основными красителями (метиленовым синим, толуидиновым синим, гематоксилином и т.д.) в виде зернистости – это скопления цистерн грЭПС. Эти органеллы отсутствуют в аксоне и в аксонном холмике, но имеются в начальных сегментах дендритов. Процесс разрушения или распада глыбок базофильного вещества называется тигролизом
и наблюдается при реактивных изменениях нейронов (например, при их повреждении) или при их дегенерации.
Хроматофильное вещество
цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при окрашивании нервных клеток основными красителями (метиленовым синим, толуидиновым синим, гематоксилином и т.д.) в виде зернистости – это скопления цистерн грЭПС. Эти органеллы отсутствуют в аксоне и в аксонном холмике, но имеются в начальных сегментах дендритов. Процесс разрушения или распада глыбок базофильного вещества называется тигролизом
и наблюдается при реактивных изменениях нейронов (например, при их повреждении) или при их дегенерации.
 Нейрофибриллы
– это цитоскелет, состоящий из нейрофиламентов и нейротубул, формирующих каркас нервной клетки. Нейрофиламенты
представляют собой промежуточные филаменты
диаметром 8-10 нм, образованные фибриллярными белками. Основной функцией этих элементов цитоскелета является опорная – для обеспечения стабильной формы нейрона. Подобную же роль играют тонкие микрофиламенты
(поперечный диаметр 6-8 нм), содержащие белки актины. В отличие от микрофиламентов в других тканях и клетках, они не соединяются с микромиозинами, что делает невозможным активные сократительные функции в зрелых нервных клетках.
Нейрофибриллы
– это цитоскелет, состоящий из нейрофиламентов и нейротубул, формирующих каркас нервной клетки. Нейрофиламенты
представляют собой промежуточные филаменты
диаметром 8-10 нм, образованные фибриллярными белками. Основной функцией этих элементов цитоскелета является опорная – для обеспечения стабильной формы нейрона. Подобную же роль играют тонкие микрофиламенты
(поперечный диаметр 6-8 нм), содержащие белки актины. В отличие от микрофиламентов в других тканях и клетках, они не соединяются с микромиозинами, что делает невозможным активные сократительные функции в зрелых нервных клетках.
Нейротубулы по основным принципам своего строения фактически не отличаются от микротрубочек. Они, как и все микротрубочки имеют поперечный диаметр около 24 нм, кольца замыкают 13 молекул глобулярного белка тубулина. В нервной ткани микротрубочки выполняют очень важную, если не сказать уникальную роль. Как и всюду они несут каркасную (опорную) функцию, обеспечивают процессы циклоза. Микротрубки полярны. Именно полярность микротрубки, в которой имеется отрицательно и положительно заряженные концы, позволяет контролировать диффузионно-транспортные потоки в аксоне (так называемый быстрый и медленный аксоток). Их подробное описание приведем ниже.
Кроме этого, в нейронах довольно часто можно видеть липидные включения (зерна липофусцина). Они характерны для старческого возраста и часто появляются при дистрофических процессах. У некоторых нейронов в норме обнаруживаются пигментные включения (например, с меланином), что обуславливает окрашивание нервных центров, содержащих подобные клетки (черная субстанция, голубоватое пятно).
Нейроны в энергетическом отношении крайне зависимы от аэробного фосфорилирования и во взрослом состоянии фактически не способы к анаэробному гликолизу. В связи с этим нервные клетки находятся в выраженной зависимости от поступления кислорода и глюкозы и при нарушении кровотока нервные клетки практически сразу прекращают свою жизнедеятельность. Момент прекращения кровотока в головном мозге означает начало клинической смерти. При мгновенной смерти, при комнатной температуре, и нормальной температуре тела процессы саморазрушения в нейронах обратимы в течение 5-7 минут. Это и является сроком клинической смерти, когда возможно оживление организма. Необратимые изменения в нервной ткани приводят к переходу от клинической смерти к биологической.
В теле нейронов можно видеть также транспортные пузырьки, часть из которых содержит медиаторы и модуляторы. Они окружены мембраной. Их размеры и строение зависят от содержания того или иного вещества.
Дендриты – короткие отростки, нередко сильно ветвятся. Дендриты в начальных сегментах содержат органеллы подобно телу нейрона. Хорошо развит цитоскелет.
Аксон (нейрит) чаще всего длинный, слабо ветвится или не ветвится. В нем отсутствует грЭПС. Микротрубочки и микрофиламенты располагаются упорядочено. В цитоплазме аксона видны митохондрии, транспортные пузырьки. Аксоны в основном миелинизированы и окружены отростками олигодендроцитов в ЦНС, или леммоцитами в периферической нервной системе. Начальный сегмент аксона нередко расширен и имеет название аксонного холмика, где происходит суммация поступающих в нервную клетку сигналов, и если возбуждающие сигналы достаточной интенсивности, то в аксоне формируется потенциал действия и возбуждение направляется вдоль аксона, передаваясь на другие клетки (потенциал действия).
Аксоток (аксоплазматический транспорт веществ). Нервные волокна имеют своеобразный структурный аппарат – микротрубочки, по которым перемещаются вещества от тела клетки на периферию (антероградный аксоток ) и от периферии к центру (ретроградный аксоток ).
Различают быстрый (со скоростью 100-1000 мм/сут.) и медленный (со скоростью 1-10 мм/сут.) аксоток. Быстрый аксоток – одинаков для различных волокон; требует значительной концентрации АТФ; происходит с участием транспортных пузырьков. Он осуществляет транспорт медиаторов и модуляторов. Медленный аксоток – за счет него от центра к периферии распространяются биологически активные вещества, а также компоненты мембран клеток и белков.
Нервный импульс передаётся по мембране нейрона в определённой последовательности: дендрит – перикарион – аксон.
Классификация нейронов
 1. По морфологии (по количеству отростков) выделяют:
1. По морфологии (по количеству отростков) выделяют:
- мультиполярные нейроны (г) — с множеством отростков (их большинство у человека),
- униполярные нейроны (а) — с одним аксоном,
- биполярные нейроны (б) — с одним аксоном и одним дендритом (сетчатка глаза, спиральный ганглий).
- ложно- (псевдо-) униполярные нейроны (в) – дендрит и аксон отходят от нейрона в виде одного отростка, а затем разделяются (в спинномозговом ганглии). Это вариант биполярных нейронов.
2. По функции (по расположению в рефлекторной дуге) выделяют:
 - афферентные (чувствительные
) нейроны (стрелка слева) – воспринимают информацию и передают ее в нервные центры. Типичными чувствительными являются ложноуниполярные и биполярные нейроны спинномозговых и черепно-мозговых узлов;
- афферентные (чувствительные
) нейроны (стрелка слева) – воспринимают информацию и передают ее в нервные центры. Типичными чувствительными являются ложноуниполярные и биполярные нейроны спинномозговых и черепно-мозговых узлов;
- ассоциативные (вставочные ) нейроны осуществляют взаимодействие между нейронами, их большинство в ЦНС;
- эфферентные (двигательные ) нейроны (стрелка справа) генерируют нервный импульс и передают возбуждение другим нейронам или клеткам других видов тканей: мышечным, секреторным клеткам.
Синапсы
Синапсы – это специфические контакты нейронов, обеспечивающие передачу возбуждения от одной нервной клетки к другой. В зависимости от способов передачи возбуждения выделяют химические и электрические синапсы.
Эволюционно более древними и примитивными являются электрические синаптические контакты . Они по строению близки к щелевидным контактам (нексусам). Считается, что обмен происходит в обе стороны, но имеются случаи, когда возбуждение передаются в одном направлении. Такие контакты часто встречаются у низших беспозвоночных и хордовых. У млекопитающих электрические контакты имеют большое значение в процессе межнейронных взаимодействий в эмбриональном периоде развития. Подобный вид контактов у взрослых млекопитающих имеет место в ограниченных участках, например их можно видеть в мезэнцефалическом ядре тройничного нерва.
Химические синапсы . Химические синапсы для передачи возбуждения от одной нервной клетки к другой используют специальные вещества – медиаторы , от чего и получили свое название. Кроме медиаторов ими используются и модуляторы . Модуляторы это специальные химические вещества, которые сами возбуждения не вызывают, но могут либо усиливать, либо ослаблять чувствительность к медиаторам (то есть модулировать пороговую чувствительность клетки к возбуждению).
Химический синапс обеспечивает однонаправленную передачу возбуждения. Строение химического синапса:
1) Пресинаптическая зона – пресинаптическое расширение, наиболее часто представляющее собой терминаль аксона, в котором содержатся синаптические пузырьки, элементы цитоскелета (нейротубулы и нейрофиламенты), митохондрии;
2) Синаптическая щель , которая принимает медиаторы из пресинаптической зоны;
3) Постсинаптическая зона – это электронноплотное вещество с рецепторами к медиатору на мембране другого нейрона.

ФИЛЬМ СИНАПСЫ
Классификация синапсов
:
1. В зависимости от того, какие структуры двух нейронов взаимодействуют в синапсе, можно выделить:
Аксо-дендритические (пресинаптическая структура аксон, постсинаптическая — дендрит);
Аксо-аксональные;
Аксо-соматические.
2. По функции выделяют:
- возбуждающие синапсы, которые приводят к деполяризации постсинаптической мембраны и активации нервной клетки;
- тормозные синапсы , которые приводят к гиперполяризации мембраны, что снижает пороговую чувствительность нейрона к внешним влияниям.
3. По основному медиатору, содержащемуся в синаптических пузырьках, синапсы делятся на группы:
- Холинергические (ацетилхолинергические): возбуждающие и тормозные;
- Адренергические (моноаминергические, норадренергические, дофаминергические): в основном, возбуждающие, но есть и тормозные;
- Серотонинергические (иногда приписываются к предыдущей группе): возбуждающие;
- ГАМК-ергические (медиатор гаммааминомаслянная кислота): тормозные;
- Пептидергические (медиаторы – большая группа вешеств, в основном: вазоинтерстициальный полипептид, вазопрессин, вещество Р (медиатор боли), нейропептид Y, окситоцин, бета-эндорфин и энкефалины (противоболевые), динорфин и т.д.).
Синаптические пузырьки отделены от гиалоплазмы одной мембраной. Холинсодержащие пузырьки электронносветлые, диаметром 40-60 мкм. Адренсодержащие – с электронноплотной сердцевиной, светлой каемкой, диаметром 50-80 мкм. Глицинсодержащие и ГАМК-содержащие – имеют овальную форму. Пептидсодержащие – с электронноплотной сердцевиной, светлой каемкой, диаметром 90-120 мкм.
Механизм передачи возбуждения в химическом синапсе: импульс, приходящий по афферентному волокну, вызывает возбуждение в пресинаптической зоне и приводит к выделению медиатора через пресинаптическую мембрану. Медиатор поступает в синаптическую щель. На постсинаптической мембране имеются рецепторы к нейромедиатору (холинорецепторы для медиатора ацетилхолина; адренорецепторы для норадреналина). В последующем связь медиаторов с рецепторами разрывается. Медиатор либо метаболизируется, либо подвергается обратному всасыванию пресинаптическими мембранами, либо захватывается мембранами астроцитов с последующей передачей медиатора к нервным клеткам.
Регенерация нейронов. Для нейронов характерна только внутриклеточная регенерация. Они являются стабильной популяцией клеток и в обычных условиях не делятся. Но имеются исключения. Так, доказана способность к делению у нервных клеток в эпителии обонятельного анализатора, в некоторых ганглиях (скоплениях нейронов вегетативной нервной системы) животных.
Нейроглия
Нейроглия
— группа клеток нервной ткани, находящиеся между нейронами, различают микроглию и макроглию
.
Макроглия
Макроглия ЦНС подразделяется на следующие клетки: астроциты (волокнистые и протоплазматические), олигодендроциты и эпендимоциты (в том числе и танициты).
Макроглия периферической нервной системы : сателлитоциты и леммоциты (шванновские клетки).
Функции макроглии: защитная, трофическая, секреторная.
Астроциты – звездчатые клетки, многочисленные отростки которых ветвятся и окружают другие структуры мозга. Астроциты есть только в ЦНС и анализаторах – производных нервной трубки.
Виды астроцитов: волокнистые и протоплазматические астроциты.
Терминали отростков обоих типов клеток имеют пуговичные расширения (ножки астроцитов), большинство из которых заканчивается в периваскулярном пространстве, окружая капилляры и образуя периваскулярные глиальные мембраны.
Волокнистые астроциты имеют многочисленные, длинные, тонкие, слабо или совсем не ветвящиеся отростки. В основном присутствуют в белом веществе мозга.
Протоплазматические астроциты отличаются короткими, толстыми и сильно ветвящимися отростками. Имеются преимущественно в сером веществе мозга. Астроциты располагаются между телами нейронов, немиелинизированной и миелинизированной частями нервных отростков, синапсами, кровеносными сосудами, подэпендимными пространствами, изолируя и в то же время структурно связывая их.
Специфическим маркером астроцитов является глиальный фибриллярный кислый белок, из которого образуются промежуточные филаменты.
Астроциты имеют относительно крупные светлые ядра, со слабо развитым ядрышковым аппаратом. Цитоплазма слабо оксифильная, в ней слабо развита аЭПС и грЭПС, комплекс Гольджи. Митохондрий мало, они небольших размеров. Цитоскелет развит умеренно в протоплазматических и хорошо – в волокнистых астроцитах. Между клетками значительное число щелевидных и десмосомоподобных контактов.
В постнатальный период жизни человека астроциты способны к миграции, особенно в зоны повреждения и способны к пролиферации (из них образуются доброкачественные опухоли астроцитомы).
Основные функции астроцитов : участие в гематоэнцефалическом и ликворогематическом барьерах (своими отростками покрывают капилляры, поверхности мозга и участвуют в транспорте веществ от сосудов к нейронам и наоборот), в связи с этим выполняют защитную, трофическую, регуляторную функции; фагоцитоз погибших нейронов, секреция биологически активных веществ: ФРФ, ангиогенные факторы, ЭФР, интерлейкин–I, простагландины.
Олигодендроциты – клетки с небольшим числом отростков, способные к образованию миелиновых оболочек вокруг тел и отростков нейронов. Олигодендроциты находятся в сером и белом веществе ЦНС, в периферической нервной системе располагаются разновидности олигодендроцитов – леммоциты (шванновские клетки). Олигодендроциты и их разновидности характеризуются способностью образовывать дупликатуру мембраны – мезаксон , который окружает отросток нейрона, образуя миелиновую или безмиелиновую оболочку.
Ядра олигодендроцитов мелкие, округлые, темноокрашенные, отростки тонкие, не ветвятся или слабо ветвятся. На электроннооптическом уровне в цитополазме хорошо развиты органеллы, особенно синтетический аппарат, слабо развит цитоскелет.
Часть олигодендроцитов концентрируется в непосредственной близости к телам нервных клеток (сателлитные, или мантийные олигодендроциты ). Терминальная зона каждого отростка участвует в формировании сегмента нервного волокна, то есть каждый олигодендроцит обеспечивает окружение сразу нескольких нервных волокон.
Леммоциты (шванновские клетки ) периферической нервной системы характеризуются удлиненными, темноокрашенными ядрами, слабо развитыми митохондриями и синтетическим аппаратом (гранулярная, гладкая ЭПС, пластинчатый комплекс). Леммоциты окружают отростки нейронов в периферической нервной системе, образуя миелиновую или безмиелиновую оболочки. В области формирования корешков спинномозговых и черепно-мозговых нервов леммоциты формируют скопления (глиальные пробки), предотвращая проникновение отростков ассоциативных нейронов ЦНС за ее пределы.
В периферической нервной системе, помимо леммоцитов, имеются другие разновидности олигодендроцитов: сателлитные (мантийные) глиоциты в периферических нервных узлах вокруг тел нейронов, глиоциты нервных окончаний , конкретные морфологические особенности которых рассматриваются при изучении нервных окончаний и анатомии нервных узлов.
Основные функции олигодендроцитов и их разновидностей : образуя миелиновую или безмиелиновую оболочки вокруг нейронов, обеспечивают изолирующей, трофической, опорной, защитной функциями; участвуют в проведении нервного импульса, в регенерации поврежденных нервных клеток, фагоцитозе остатков осевых цилиндров и миелина при нарушении структуры аксона дистальнее места повреждения.
Эпендимоциты , или эпендимная глия – клетки низкопризматической формы, образующие непрерывный пласт, покрывающий полости мозга. Эпендимоциты тесно прилежат друг к другу, формируя плотные, щелевидные и десмосомальные контакты. Апикальная поверхность содержит реснички, которые у большинства клеток затем замещаются микроворсинками. Базальная поверхность имеет базальные впячивания (инвагинации), а также длинные тонкие отростки (от одного до нескольких), которые проникают до периваскулярных пространств микрососудов мозга.
В цитоплазме эпендимоцитов обнаруживаются митохондрии, умеренно развитый синтетический аппарат, хорошо представлен цитоскелет, имеется значительное количество трофических и секреторных включений.
Вариантом эпендимной глии являются танициты . Они выстилают сосудистые сплетения желудочков головного мозга, субкомиссуральный орган задней комиссуры. Активно участвуют в образовании ликвора (спинномозговой жидкости). Характеризуются тем, что базальная часть содержит тонкие длинные отростки.
Основные функции эпендимоцитов : секреторная (синтез ликвора), защитная (обеспечение гемато-ликворного барьера ), опорная, регуляторная (предшественники таницитов направляют миграцию нейробластов в нервной трубке в эмбриональном периоде развития).
Микроглия
Микроглиоциты, или нейральные макрофаги – клетки небольших размеров мезенхимного происхождения (производные моноцитов), диффузно распределенные в ЦНС, с многочисленными сильно ветвящимися отростками, способны к миграции. Микроглиоциты – специализированные макрофаги нервной системы. Их ядра характеризуются преобладанием гетерохроматина. В цитоплазме обнаруживается много лизосом, гранул липофусцина; синтетический аппарат развит умеренно.
Функции микроглии: защитная (в том числе иммунная).
Нервные волокна
Нервное волокно состоит из отростка нейрона – осевого цилиндра (дендрита или аксона) и оболочки олигодендроцита или его разновидностей .
Виды нервных волокон:
1) В зависимости от того, как произошло образование оболочки, нервные волокна подразделяются на миелиновые и безмиелиновые.
В периферической нервной системе нервные волокна окружают леммоциты. Один леммоцит связан с одним нервным волокном. В центральной нервной системе отростки нейронов окружают олигодендроциты. Каждый олигодендроцит участвует в формировании нескольких нервных волокон.
Миелинизация волокон осуществляется путем удлинения и «наворачивания» мезаксона вокруг отростка нервной клетки (в периферической нервной системе) или удлинения и вращения отростка олигодендроцита вокруг осевого цилиндра в ЦНС.
Миелиновые (мякотные) волокна в периферической нервной системе имеют в своём составе один отросток нейрона, окружённый удлинённой дупликутурой леммоцита (мезаксон). В миелиновом волокне мезаксон многократно оборачивается вокруг осевого цилиндра, формируя многократные витки мембраны – миелин. Зоны разрыхления миелина (проникновения цитоплазмы леммоцита) называются насечками (Шмидта-Лантермана). Каждый леммоцит образует сегмент волокна, участки границ соседних клеток немиелинизированы и называются перехватами Ранвье , таким образом, по длине волокна миелиновая оболочка имеет прерывистый ход. Миелиновая оболочка является биологическим изолятором. Распространение деполяризации в миелиновом волокне осуществляется скачками от перехвата к перехвату.
Безмиелиновые (безмякотные) волокна в периферической нервной системе состоят из одного или нескольких осевых цилиндров, погружённых в цитолемму окружающего их леммоцита. Мезаксон (дупликатура мембраны) короткий. Передача возбуждения в безмиелиновых волокнах происходит по поверхности нерва через изменение поверхностного заряда.
2) В зависимости от скорости проведения нервного импульса различают следующие типы нервных волокон:
- Тип А имеет подгруппы:
- А a — обладают наибольшей скоростью проведения возбуждения — 70-120 м/с (соматические двигательные нервные волокна);
- А b — скорость проведения составляет 40-70 м/с. Это соматические афферентные нервы и некоторые эфферентные соматические нервы;
- А g — скорость проведения составляет 15-40 м/с — афферентные и эфферентные симпатические и парасимпатические нервы;
- А d (дельта) — скорость проведения 5-18 м/с. По этой группе афферентных соматических нервов проводятся первичная (быстрая) боль.
- Тип В – скорость проведения от 3 до 14 м/с – преганглионарные симпатические волокна, некоторые парасимпатические волокна, то есть это вегетативные нервы.
- Тип С – скорость проведения 0,5-3 м/с: постганглионарные вегетативные волокна (безмиелиновые). Проводят болевые импульсы медленной вторичной боли (от рецепторов пульпы зуба).
Нейрогенез. На 15-17 сутки внутриутробного развития человека под индуцирующим влиянием хорды из первичной эктодермы формируется нервная пластинка (скопление продольно лежащего клеточного материала). С 17 по 21 сутки пластинка инвагинирует и превращается сначала в нервный желобок , а затем в трубку . К 25 суткам эмбриогенеза происходит отщепление нервной трубки от эктодермы и замыкание переднего и заднего отверстий (нейропоров). По бокам от нервного желобка располагаются структуры нервного гребня .
На ранних сроках развития нервная трубка сформирована медулобластами – стволовыми клетками нервной ткани ЦНС. Из нервного гребня образуется ганглиозная пластинка состоящая из ганглиобластов – стволовых клеток нейронов и нейроглии периферической нервной системы. Медулобласты и ганглиобласты интенсивно иммигрируют, делятся и затем дифференцируются.
В ранние сроки внутриутробного развития нервная трубка представляет собой пласт отростчатых клеток, лежащих в виде одного слоя, но в несколько рядов. Изнутри и снаружи они ограничены пограничными мембранами. На внутренней поверхности (прилежащей к полости нервной трубки) медулобласты делятся.
В последующем нервная трубка формирует несколько слоев . Среди них можно выделить:
- Внутренняя пограничная мембрана : отделяет полость нервной трубки от клеток;
- Эпендимный слой (вентрикулярный в области мозговых пузырей) представлен бластными клетками-предшественниками макроглии;
- Субвентрикулярная зона (только в передних мозговых пузырях), где происходит пролиферация нейробластов;
- Мантийный (плащевой) слой , содержащий мигрирующие и дифференцирующиеся нейробласты и глиобласты;
- Маргинальный слой (краевая вуаль) сформирован отростками глиобластов и нейробластов. В ней можно видеть тела отдельных клеток.
- Наружная пограничная мембрана .
Диффероны нервной ткани центральной нервной системы
- Дифферон нейрона: медулобласт – нейробласт – молодой нейрон – зрелый нейрон.
- Дифферон астроцита: медулобласт – спонгиобласт – астробласт – протоплазматический или волокнистый астроцит.
- Диферрон олигодендроцита: медулобласт — спонгиобласт – олигодендробласт – олигодендроцит.
- Дифферон эпендимной глии: медулобаст – эпендимобласт – эпендимоцит или таницит.
- Дифферон микроглии: стволовая клетка крови – полустволовая клетка крови (КОЕ ГЭММ) – КОЕ ГМ – КОЕ М – монобласт – промоноцит – моноцит – микроглиоцит покоя – активированный микроглиоцит.
Диффероны нервной ткани в периферической нервной системе
1. Дифферон нейрона: ганглиобласт – нейробласт – молодой нейрон – зрелый нейрон.
2.Дифферон леммоцита: ганглиобласт – глиобласт – леммоцит (шванновская клетка).
Механизмы нейрогенеза. В процессе внутриутробного развития нейробласты мигрируют в области анатомических закладок нервных центров. При этом они прекращают делиться. В ЦНС миграция нейробластов контролируется адгезивными межклеточными взаимодействиями (с помощью кадгеринов и интегринов радиальной глии), сигнальными молекулами межклеточного вещества (в том числе фибронектинами и ламининами). После того как нейробласты достигают области своей постоянной локализации, они начинают дифференцироваться и формировать отростки. Направление роста отростков также контролируется упомянутыми адгезивными молекулами (кадгерины, интегрины, сигнальные молекулы межклеточного вещества).
Во внутриутробном развитии и после рождения происходит конкурентное взаимодействие между аналогичными нейронами нервных центров. При этом нервные клетки, не успевшие занять соответствующую зону, либо сформировать контакты, подвергаются апоптозу. В раннем развитии погибает от трети до половины нервных клеток.
В последующем развитии вокруг нервных клеток формируется глиальное окружение и происходит миелинизация нервных волокон. Нервные клетки до полового созревания продолжают формировать отростки и синаптические контакты. Максимального развития нервная ткань достигает к 25-30 годам.
С возрастом наблюдается гибель части нервных клеток и компенсаторная гипертрофия других. В нейронах может накапливаться липофусцин. Области с погибшими телами нервных клеток замещаются глиальными рубцам, образованными скоплением гипертрофированных астроцитов.
Дендриты сильно ветвятся, образуя дендритное дерево, и обычно короче аксона. От дендритов возбуждение направляется к телу нервной клетки. Они формируют постсинаптические структуры, воспринимающие возбуждение. Дендритов много, но может быть один. Аксон присутствует всегда, по одному на каждую нервную клетку. Он не ветвится или слабо ветвится в терминальных областях и заканчивается синаптическим бутоном, передающим возбуждение на другие клетки (пресинаптическая зона). Нейроны передают возбуждение с помощью специализированных контактов (синапсов). Вещество, обеспечивающее передачу возбуждения, называется медиатором . В каждом нейроне обычно обнаруживается один основной медиатор.
Регенерация нервных волокон в периферической нервной системе
После перерезки нервного волокна проксимальная часть аксона подвергается восходящей дегенерации, миелиновая оболочка в области повреждения распадается, перикарион нейрона набухает, ядро смещается к периферии, хроматофильная субстанция распадается. Дистальная часть, связанная с иннервируемым органом, претерпевает нисходящую дегенерацию с полным разрушением аксона, распадом миелиновой оболочки и фагоцитозом детрита макрофагами и глией. Леммоциты сохраняются и митотически делятся, формируя тяжи – ленты Бюнгнера. Через 4-6 недель структура и функция нейрона восстанавливается, от проксимальной части аксона дистально отрастают тонкие веточки, растущие вдоль лент Бюнгнера. А результате регенерации нервного волокна восстанавливается связь с органом-мишенью. При возникновении преграды на пути регенерирующего аксона (например, соединительнотканного рубца), восстановления иннервации не происходит.
С дополнениями из учебно-методического пособия «Общая гистология» (составители: Шумихина Г.В., Васильев Ю.Г., Соловьёв А.А., Кузнецова В.М., Соболевский С.А., Игонина С.В., Титова И.В., Глушкова Т.Г.)
Частная гистология.
Частная гистология – наука о микроскопическом строении и происхождении органов. Каждый орган состоит из 4 тканей.
Органы нервной системы.
По функциональному признаку
1. Соматическую нервную систему – участвует в иннервации тела человека и высшей нервной деятельности.
a. Центральный отдел :
i. Спинной мозг – ядра задних и передних рогов
ii. Головной мозг – кора мозжечка и больших полушарий
b. Периферический отдел :
i. Спинномозговые ганглии
ii. Черепномозговые ганглии
iii. Нервные стволы
2. Вегетативную нервную систему – обеспечивает работу внутренних органов, иннервирует гладкие миоциты и представляет секреторные нервы.
1) Симпатическая :
a. Центральный отдел :
i. Спинной мозг – ядра боковых рогов торако-люмбального отдела
ii. Головной мозг – гипоталамус
b. Периферический отдел :
i. Симпатические ганглии
ii. Нервные стволы
2) Парасимпатическая :
a. Центральный отдел :
i. Спинной мозг – ядра боковых рогов сакрального отдела
ii. Головной мозг – ядра ствола, гипоталамус
b. Периферический отдел :
i. Парасимпатические ганглии
ii. Нервные стволы
iii. Спинномозговые и черепно-мозговые ганглии
По анатомическому признаку органы нервной системы делятся на:
1. Периферическую нервную систему.
2. Центральную нервную систему.
Эмбриональные источники развития :
1. Нейроэктодерма (даёт начало паренхиме органов).
2. Мезенхима (даёт начало строме органов, совокупности вспомогательных структур, обеспечивающих функционирование паренхимы).
Органы нервной системы функционируют в относительной изоляции от окружающей среды, отделяясь от неё биологическими барьерами . Виды биологических барьеров :
1. Гематонейральный (отграничивает кровь от нейронов).
2. Ликворонейральный (отграничивает ликвор от нейронов).
3. Гематоликворный (отграничивает ликвор от крови).
Функции нервной системы:
1. Регуляция функций отдельных внутренних органов.
2. Интеграция внутренних органов в системы органов.
3. Обеспечение взаимосвязи организма с внешней средой.
4. Обеспечение высшей нервной деятельности.
Все функции основываются на принципе рефлекса . Материальной основой является рефлекторная дуга , состоящая из 3-х звеньев: афферентного , ассоциативного и эфферентного . Они распределены по отдельным органам нервной системы.
Органы периферической нервной системы :
1. Нервные стволы (нервы).
2. Нервные узлы (ганглии).
3. Нервные окончания.
Нервные стволы – это пучки нервных волокон, объединённые системой соединительно-тканных оболочек. Нервные стволы смешаны, т.е. в каждом есть миелиновые и амиелиновые волокна, в результате чего обслуживается соматическая и вегетативная нервные системы.
Строение нервного ствола :
1. Паренхима : безмиелиновые и миелиновые нервные волокна + микроганглии.
2. Строма : соединительно-тканные оболочки:
1) Периневрий (периневральные влагалища: РВНСТ + кровеносные сосуды + эпендимоглиоциты + цереброспинальная жидкость).
2) Эпиневрий (ПВНСТ + кровеносные сосуды).
3) Периневрий (отщепление от эпиневрия внутрь ствола).
4) Эндоневрий (РВНСТ + кровеносные сосуды).
В периневрии есть щелевидное пространство – щелевидное периневральное влагалище , которое заполнено ликвором (циркулирующей биологической жидкостью). Структурные компоненты стенок периневрального влагалища :
1. Низкопризматические эпендимоглиоциты.
2. Базальная мембрана.
3. Субэпендимальная пластинка.
4. Кровеносные сосуды.
Ликвор в периневральном влагалище может отсутствовать. В них иногда вводят анестезирующие вещества, антибиотики (т.к. по ним распространяется заболевание).
Функции нервных стволов :
1. Проводниковая (проводят нервный импульс).
2. Трофическая (питательная).
4. Являются начальным звеном в секреции и циркуляции цереброспинальной жидкости.
Регенерация нервных стволов :
1. Физиологическая регенерация (очень активное восстановление оболочек за счёт фибробластов).
2. Репаративная регенерация (восстанавливается тот участок нервного ствола, нервные волокна которого не утратили связи с перикарионом – они способны расти на 1мм/сутки; периферические отрезки нервных волокон не восстанавливаются).
Нервные узлы (ганглии) – группы или кооперации нейронов, вынесенные за пределы мозга. Нервные узлы «одеты» в капсулы.
Типы ганглиев:
1. Спинномозговые .
2. Черепномозговые .
3. Вегетативные .
Спинномозговые ганглии – утолщения на начальных отделах задних корешков спинного мозга; это скопление афферентных (чувствительных) нейронов (они являются первыми нейронами в цепи рефлекторной дуги).
Строение спинномозгового ганглия :
1. Строма :
1) наружная соединительно-тканная капсула, состоящая из 2 листков:
a. наружный листок (плотная соединительная ткань – продолжение эпиневрия спинномозгового нерва)
b. внутренний листок (многотканевой: РВНСТ, глиоциты; аналог периневрия спинномозгового нерва; есть расщепления, переходящие на внутриорганные перегородки, заполнены ликвором).
2) внутриорганные перегородки, отходящие от капсулы внутрь узла
b. кровеносные и лимфатические сосуды
c. нервные волокна
d. нервные окончания
3) собственные соединительно-тканные капсулы псевдоуниполярных нейронов
a. волокнистая соединительная ткань
b. однослойный плоский эпендимоглиальный эпителий
c. перинейрональное пространство с цереброспинальной жидкостью
2. Паренхима :
1) центральная часть (миелиновые нервные волокна – отростки псевдоуниполярных нейронов)
2) периферическая часть (псевдоуниполярные нейроны + мантийные глиоциты (олигодендроглиоциты)).
Функции спинномозгового ганглия:
1. Участие в рефлекторной деятельности (первые нейроны в цепи рефлекторной дуги).
2. Являются начальным звеном в обработке афферентной информации.
3. Барьерная функция (гематонейральный барьер).
4. Являются звеном в циркуляции ликвора.
Источники эмбрионального развития спинномозгового ганглия :
1. Ганглиозная пластинка (даёт начало элементам паренхимы органа).
2. Мезенхима (даёт начало элементам стромы органа).
Ганглии вегетативной нервной системы – расположены после спинного мозга, участвуют в создании вегетативных дуг.
Типы ганглиев вегетативной нервной системы :
1. Симпатические :
1) Паравертебральные;
2) Превертебральные;
2. Парасимпатические :
1) Внутриорганные (интрамуральные);
2) Околоорганные (параорганные);
3) Вегетативные узлы головы (по ходу черепно-мозговых нервов).
Строение ганглиев вегетативной нервной системы :
1. Строма : строение аналогично строме спинномозгового ганглия.
2.1. Паренхима симпатических ганглиев : нейроны, расположенные хаотично по всему ганглию + клетки-сателлиты + соединительно-тканная капсула.
1) крупные длинноаксонные мультиполярные эфферентные адренергические нейроны
2) мелкие равноотростчатые мультиполярные ассоциативные адренергические интенсивно флюоресцирующие (МИФ) – нейроны
3) преганглионарные миелиновые холинэргические волокна (аксоны нейронов боковых рогов спинного мозга)
4) постганглионарные безмиелиновые адренергические нервные волокна (аксоны крупных нейронов ганглиев)
5) внутриганглионарные безмиелиновые ассоциативные нервные волокна (аксоны МИФ – нейронов).
2.2. Паренхима парасимпатических ганглиев :
1) длинноаксонные мультиполярные эфферентные холинэргические нейроны (Догеля I типа).
2) длиннодендритные мультиполярные афферентные холинэргические нейроны (Догеля II типа): дендрит – к рецептору, аксон – к 1 и 3 типу.
3) равноотростчатые мультиполярные ассоциативные холинэргические нейроны (Догеля III типа).
4) преганглионарные миелиновые холинергические нервные волокна (аксоны боковых рогов спинного мозга).
5) постганглионарные безмиелиновые холинергические нервные волокна (аксоны нейронов Догеля I типа).
Функции ганглиев вегетативной нервной системы :
1. Симпатических :
1) Проведение импульсов к рабочим органам (2.1.1)
2) Распространение импульса в пределах ганглия (тормозной эффект) (2.1.2)
2. Парасимпатических :
1) Проведение импульса к рабочим органам (2.2.1)
2) Проведение импульса от интерорецепторов в пределах местных рефлекторных дуг (2.2.2)
3) распространение импульса в пределах или между ганглиями (2.2.3).
Источники эмбрионального развития ганглиев вегетативной нервной системы :
1. Ганглиозная пластинка (нейроны и нейроглия).
2. Мезенхима (соединительная ткань, сосуды).
На ранних этапах развития человеческого зародыша из клеток эктодермы возникает нервная пластинка, образованная однослойным однорядным призматическим эпителием (нейроэпителием), под которым располагается хорда, индуцирующая возникновение нервной пластинки (рис. 224). Нервная пластинка быстро растет, утолщается, становится многослойной, углубляется, образуя желобок, края которого приподнимаются и превращаются в нервные валики. Под валиками формируются нервные гребни - выросты в виде тяжей клеток, которые после замыкания желобка в нервную трубку превращаются в ганглиозные пластинки, располагающиеся сбоку от нервной трубки и отделяющиеся от нее. Нервная трубка также отделяется от эктодермы. После образования трубки клетки нейроэпителия дифференцируются в субвентрикулярные нервные клетки - нейробласты, число которых быстро увеличивается благодаря активной пролиферации. Из этих клеток образуется мантийный слой. Из этих же клеток возникают и первичные опорные клетки - глиобласты, которые мигрируют в мантийный слой. Впоследствии из мантийного слоя образуется серое вещество мозга. Митотическое деление нейробластов заканчивается до формирования отростков. Вначале начинается рост аксона, позднее - дендритов. Отростки нейробластов образуют на периферии нервной трубки краевой (маргинальный) слой, из которого образуется белое вещество. Вентрикулярные клетки, расположенные на внутренней поверхности нервной трубки, дифференцируются в танициты и эпителиоидные эпендимоциты. На стадии нервной трубки ганглиозные пластинки фрагментируются, образуя округлые структуры, из которых формируются спинномозговые узлы.
Итак, три слоя стенки нервной трубки дают начало эпендиме, выстилающей полости центральной нервной системы (внутренний), серому веществу (средний, плащевой) и белому веществу (наружный) (табл. 38). Боковые отделы трубки растут более интенсивно, из их вентральных отделов возникают передние столбы серого вещества (тела клеток и волокна) и прилежащее белое вещество (только нервные волокна). Из дорсальных отделов нервной трубки образуются задние столбы серого вещества и белое вещество спинного мозга. Головной отдел нервной трубки растет неравномерно. В некоторых участках она толще, благодаря усиленному продольному росту она изгибается. Уже на 4-й неделе эмбрионального развития различают три первичных мозговых пузыря: передний, средний и задний. К концу 4-й недели передний мозговой пузырь начинает делиться на два: конечный мозг, из которого впоследствии развивается вся кора полушарий большого мозга, и промежуточный, из которого развиваются таламус и гипоталамус. Просвет трубки переднего мозга образует боковые и III желудочки. Задний (ромбовидный пузырь) в течение 5-й недели также делится на два пузыря, из которых образуются мозжечок, продолговатый мозг и мост. Из среднего пузыря, сохраняющего трубчатую форму, образуется средний мозг, просвет трубки - мозговой (сильвиев) водопровод. В результате будущий головной мозг состоит из пяти пузырей (рис. 225). В области среднего мозгового пузыря формируются ножки мозга и пластинка крыши среднего мозга. Растут боковые стенки промежуточного мозга, образуя таламусы, выросты боковых стенок дают начало глазным пузырькам. Нижняя стенка промежуточного мозга выпячивается, образуя серый бугор, воронку, подбугорье (гипоталамус) и заднюю долю гипофиза. Происхождение различных отделов мозга представлено в табл. 39 .
Важные преобразования происходят в конечном мозге. На I стадии формируются обонятельные структуры и лимбическая система (палеокортекс), расположенная вокруг краев развивающегося конечного мозга; на II стадии стенки переднего мозга утолщаются благодаря интенсивной пролиферации нейробластов, возникают зачатки базальных ганглиев; наконец, на III стадии формируется кора полушарий большого мозга (неокортекс). В связи с активным митотическим делением нейробластов неокортекса, когда скорость образования клеток достигает 250 000 в 1 мин, начинается формирование мозговых борозд и извилин полушарий большого мозга. Масса головного мозга новорожденного ребенка относительно велика, она составляет в среднем 390 г (340 - 430) у мальчиков и 355 г (330 - 370) у девочек (12 - 13 % массы тела, у взрослого человека - около 2,5%). Отношение массы мозга новорожденного к массе его тела в пять раз больше, чем у взрослого, соответственно 1: 8 и 1:40. В течение первого года жизни масса мозга удваивается, а к 3 . 4 годам утраивается, затем она медленно увеличивается и к 20 - 29 годам достигает максимальных цифр (1355 г у мужчин и 1220 г у женщин). К 20 - 25 годам и в последующем, вплоть до 60 лет у мужчин и 55 лет у женщин, масса мозга существенно не изменяется, после 55 - 60 лет она несколько уменьшается. До 4 лет жизни головной мозг ребенка растет равномерно в высоту, длину и ширину, в дальнейшем преобладает рост мозга в высоту. Наиболее быстро растут лобная и теменная доли.
У новорожденного ребенка лучше развиты филогенетически более старые отделы мозга. Масса ствола мозга равна 10 - 10,5 г (около 2,7% массы тела, у взрослого человека - около 2%). К моменту рождения ребенка продолговатый мозг, мост и их ядра развиты хорошо, масса первого составляет около 4 - 5 г, второго -3,5 - 4 г. Хуже развит мозжечок, особенно его полушария, лучше - червь, слабо выражены извилины и борозды полушарий мозжечка. Масса мозжечка новорожденного ребенка не превышает 20 г (5,4% массы тела, у взрослого - 10%). В течение первых 5 месяцев жизни масса мозжечка увеличивается в три раза, в 9 месяцев, когда ребенок умеет стоять и начинает ходить, . в четыре раза. Наиболее интенсивно развиваются полушария мозжечка. Промежуточный мозг у новорожденного развит также относительно хорошо. Формирование борозд и извилин начинается у плода начиная с 5-го месяца развития. У 7- месячного плода уже заметны борозды и извилины, к моменту рождения они развиты полностью (Ф. И. Валькер, 1951), однако ветви основных борозд и мелкие извилины выражены слабо. Формирование рельефа полушарий продолжается в течение первых 6-7 лет жизни, борозды становятся глубже, извилины между ними -рельефнее (В. В. Бунак, 1936). У новорожденного ребенка наиболее развиты височные доли и обонятельный мозг, слабее - лобные. У новорожденного ребенка кора полушарий большого мозга не полностью дифференцирована. Желудочки мозга новорожденного ребенка относительно крупнее, чем у взрослого человека. Твердая оболочка головного мозга новорожденного ребенка тонкая, плотно сращена с костями черепа, ее отростки развиты слабо. Синусы тонкостенные, относительно широкие. После 10 лет строение и топография синусов такие же, как у взрослого. Паутинная и мягкая оболочки головного и спинного мозга у новорожденного тонкие, нежные. Подпаутинное пространство относительно широкое.
Основные этапы развития мозга в эмбриогенезе были описаны еще в прошлом веке, однако до сих пор сравнительно мало известно о процессах, которые обеспечивают формирование отдельных структур мозга и их связей друг с другом.
Эмбриогенез (внутриутробное развитие) человека закономерно связан с процессами его предшествующей эволюции. Связь между ними настолько ощутима, что существует даже понятие филоэмбриогенез, подчеркивающее единство процессов эволюционного и индивидуального развития.
Онтогенетическое развитие нервной системы (греч. «onthos» - особь, сущее), то есть развитие индивидуальное, протекающее с момента оплодотворения яйцеклетки и до смерти данного индивида, в основных чертах отражает филогенез нервной системы данного вида.
Образовавшаяся после оплодотворения зигота начинает делиться и образует морулу, представляющую собой скопление клеток, способных к дифференциации в разных направлениях. Эти клетки в последующем делятся неравномерно и образуют бластулу, состоящую из трофобласта и эмбриобласта.
Из клеток наружной части эмбриобласта формируется зародышевый, или эмбриональный диск, который вскоре делится на два листка (слоя) - энтодерму (внутренний листок) и эктодерму (наружный листок). Через некоторое время между ними образуется мезодерма (средний листок). Из эктодермы в последующем образуются нервная ткань, хорда и кожа. Из клеток
энтодермы сформируются дыхательная и пищеварительная трубки, а из мезодермы будут образовываться мышцы, соединительная ткань, клетки крови, мочеполовая система и части большинства внутренних органов.
Зародышевый диск по мере роста увеличивается в длину и превращается в эмбриональную пластинку (полоску). Одновременно с этим увеличивается и толщина эмбриона.
На следующей стадии эмбрионального развития происходит сворачивание эмбриональной пластинки в зародышевую трубку. При этом энтодерма и мезодерма вворачиваются внутрь эктодермы, и образуется гаструла. На поверхности эмбриона остаётся нервная ткань в виде продольной нервной пластинки и та часть эктодермы, из которой в последующем образуется кожа.
В первичной нервной пластинке клетки-предшественницы нервной ткани изначально располагаются в один слой. Каждый сегмент этой пластинки отвечает за образование специфических структур нервной системы, хотя на очень ранних стадиях эмбриогенеза предназначение участка для формирования определенных частей мозга может меняться. Если в это время удалить некоторые участки нервной пластинки, оставшиеся ткани нервной пластинки заменят утраченные и в результате разовьётся полноценный мозг. На более поздних стадиях развития замещения не происходит, и мозг формируется не полностью.
Нервная пластинка быстро растёт, на 3-й неделе развития её края начинают утолщаться и приподниматься над
первоначальной зародышевой пластинкой. На 19-й день левый и правый края сближаются и срастаются по средней линии,образуя полую нервную трубку, располагающуюся под поверхностью эктодермы, но отдельно от нее. Процесс замыкания нервной трубки начинается на уровне 5-го сегмента, распространяясь как в головном, так и каудальном направлении.
К 25-ому дню он заканчивается. Каудальный конец нервной трубки временно смыкается с задней кишкой, образуя нейроэнтеральный канал. Клетки нервной трубки (медуллобласты) в последующем дифференцируются в нейроны головного и спинного мозга, а также в нейроглиальные клетки (олигодендроциты, астроциты и эпендимные клетки).
Во время сворачивания нервной трубки некоторые клетки нервной пластинки остаются за её пределами, и из них формируется нервный гребень. Он лежит между нервной трубкой и кожей, и в дальнейшем из клеток нервного гребня развиваются нейроны периферической нервной системы, шванновские клетки, клетки мозгового вещества надпочечников и мягкой мозговой оболочки.
Вскоре после формирования нервной трубки, тот её конец, из которого впоследствии образуется голова, закрывается.
Затем передняя часть нервной трубки начинает раздуваться, и образуются три вздутия - так называемые первичные мозговые пузыри. Одновременно с формированием этих пузырей происходит формирование в сагиттальной плоскости двух изгибов будущего головного мозга. Головной, или теменной изгиб формируется в области среднего пузыря.
Шейный изгиб отделяет зачаток головного мозга от остальной части нервной трубки, из которой в последующем сформируется спинной мозг.
Из первичных мозговых пузырей формируются три основных отдела головного мозга: передний (prosencephalon - передний мозг), средний (mesencephalon - средний мозг) и задний (rhombencephalon - задний, или ромбовидный мозг). Этот этап развития мозга называется стадией трёх мозговых пузырей. После образования трёх первичных пузырей с замыканием заднего конца нервной трубки на боковых поверхностях переднего пузыря появляются зрительные пузырьки, из которых сформируются сетчатка глаза и зрительные нервы.
Следующим этапом развития мозга является параллельное дальнейшее формирование изгибов мозговой трубки и образование из первичных пузырей пяти вторичных мозговых пузырей (стадия пяти мозговых пузырей). Первый и второй вторичные мозговые пузыри образуются за счёт деления на две части переднего первичного пузыря. Из этих пузырей в последующемформируются, соответственно, конечный мозг (полушария мозга) и промежуточный мозг. Третий вторичный мозговой пузырь образуется из неделящегося среднего первичного пузыря. Четвёртый и пятый мозговые пузыри образуются в результате деления третьего (заднего) первичного пузыря на верхнюю и нижнюю части. Из них в последующем образуется
собственно задний мозг (мозжечок и мост) и продолговатый мозг.
Всего в процессе онтогенеза мозговая трубка изгибается три раза в сагиттальной плоскости. Сначала в области среднего мозгового пузыря, рядом с образующимся перешейком мозга, отделяющим передний и средний мозг, формируется выпуклый в дорсальную сторону головной, или теменной, изгиб. Затем на границе с зачатком спинного мозга образуется шейный изгиб, также выпуклый дорсально. Третий, мостовой изгиб формируется в области заднего первичного пузыря, его выпуклая сторона обращена вперед (вентрально). Именно этот изгиб делит задний мозг на 4 и 5 вторичные пузыри.
Таким образом, после деления первичных мозговых пузырей и формирования мозговых изгибов в зачатке мозга человека дифференцируется 5 отделов, из которых в дальнейшем образуются: 1. Конечный мозг (telencephalon), 2. Промежуточныймозг (diencephalon), 3. Средний мозг (mesencephalon), 4. Задний мозг (metencephalon) и 5. Продолговатый мозг
(myelencephalon seu medula oblongata).
По мере роста нервной трубки происходит утолщение её стенок и усложнение рельефа поверхности мозговых пузырей.
Это приводит к неравномерному сужению полости нервной трубки. В результате просвет спинного мозга превращается в узкий центральный канал спинного мозга, а полости мозговых пузырей приобретают форму различных по величине и положению щелей, называемых желудочками головного мозга. Все желудочки мозга последовательно соединяются между собой и с центральным каналом спинного мозга. Они заполнены спинномозговой жидкостью, которая образуется внутрижелудочковыми сосудистыми сплетениями и эпендимными клетками. Через отверстия в нижнем мозговом парусе
спинномозговая жидкость оттекает из системы желудочков мозга в подпаутинное пространство.
По мере своего роста полушария большого мозга сначала увеличиваются в области лобной доли, затем теменной и, наконец, височной доли. Благодаря этому создаётся впечатление, будто кора мозга (плащ) вращается вокруг таламуса сначала спереди назад, затем вниз и, наконец, загибается вперёд к лобной доле. В результате к моменту рождения плащ мозга покрывает не только таламусы, но и дорсальную поверхность среднего мозга и мозжечка.
Похожая информация.
3.1.1. закладка нервной системы
Центральный и периферический отделы нервной системы человека развиваются из единого эмбрионального источника эктодермы. В процессе развития зародыша она закладывается в виде так называемой нервной пластинки группы высоких, быстро размножающихся клеток по средней линии зародыша. На. 3-й неделе развития нервная пластинка погружается в подлежащую ткань, принимает форму желобка, края которого несколько приподнимаются над уровнем эктодермы в виде нервных валиков. По мере роста зародыша нервный желобок удлиняется и достигает каудального конца зародыша. На 19-й день развития начинается процесс смыкания нервных валиков над желобком, в результате чего образуется длинная полая трубка нервная трубка, располагающаяся непосредственно под поверхностью эктодермы, но отдельно от последней.
При замыкании нервного желобка в трубку и срастании его краев материал нервных валиков оказывается зажатым между нервной трубкой и смыкающейся над ней кожной эктодермой. При этом клетки нервных валиков перераспределяются в один слой, образуя ганглиозную пластинку зачаток с очень широкими потенциями развития. Из этого эмбрионального зачатка образуются все нервные узлы соматической периферической и вегетативной нервных систем, включая внутриорганные нервные элементы.
Процесс замыкания нервной трубки начинается на уровне 5-го сегмента, распространяясь как в головном, так и в каудальном направлении. К 24-му дню развития он заканчивается в головной части, сутками позже в каудальной. Каудальный конец нервной трубки временно смыкается с задней кишкой, образуя нейроэнтеральный канал.
Сформировавшаяся нервная трубка в головном конце, на месте образования будущего головного мозга, расширяется. Более тонкая каудальная часть ее преобразуется в спинной мозг.
Параллельно с формированием нервной трубки происходит образование других структур (хорда, мезодерма), которые вместе с нервной трубкой составляют так называемый комплекс осевых зачатков. С образованием комплекса осевых зачатков зародыш человека, лишенный ранее оси симметрии, приобретает билатеральную симметрию. Теперь уже совершенно отчетливо в нем различимы головной и каудальный отделы, правая и левая половины туловища.
Развитие различных отделов центральной и периферической нервных систем в преи постнатальном онтогенезе человека происходит неравномерно. Особенно сложный путь развития проходит центральная нервная система.
Клетки сформированной нервной трубки, которые в своем дальнейшем развитии дадут начало как нейронам, так и глиоцитам, носят название медуллобластов. Клеточные элементы ганглиозной пластинки, обладающие, по-видимому, такими же гистогенетическими потенциями, называются ганглиобластами. Следует отметить, что на начальных этапах дифференцировки нервной трубки и ганглиозной пластинки их клеточный состав однороден.
В своей дальнейшей дифференцировке медуллобласты детерминируются частью в нейтральном направлении, превращаясь в нейробласты, частью в нейроглиальном направлении, образуя спонгиобласты.
Нейробласты отличаются от нейронов значительно меньшей величиной, отсутствием дендритов и синаптических связей (следовательно, они не включены в рефлекторные дуги), а также отсутствием субстанции Ниссля в цитоплазме. Однако они уже имеют слабовыраженный нейрофибриллярный аппарат, формирующийся аксон и характеризуются отсутствием способности к митотическому делению.
В спинальном отделе первичная нервная трубка рано делится на три слоя: внутренний эпендимный, промежуточный мантийный (или плащевой) и наружный светлый краевую вуаль.
Эпендимный слой дает начало нейронам и глиальным клеткам (эпендимоглии) центральной нервной системы. В его составе обнаруживаются нейробласты, которые впоследствии мигрируют в мантийный слой. Остающиеся в эпендимном слое клетки прикрепляются к внутренней пограничной мембране, посылают наружу отростки, участвуя тем самым в образовании наружной пограничной мембраны. Они носят название спонгиобластов, которые в случае утери связи с внутренней и наружной пограничными мембранами превратятся в астроцитобласты. Те клетки, которые сохранят свою связь с внутренней и наружной пограничными мембранами, превратятся в эпендимные глиоциты, выстилающие у взрослого центральный канал спинного мозга и полости желудочков мозга. Они приобретают в процессе дифференцировки реснички, способствующие току цереброспинальной жидкости.
Эпендимный слой нервной трубки как в туловищном, так и в головном ее отделе сохраняет до сравнительно поздних стадий эмбриогенеза потенцию к образованию весьма разнообразных тканевых элементов нервной системы.
В мантийном слое развивающейся нервной трубки располагаются нейробласты и спонгиобласты, дающие при дальнейшей дифференцировке астроглию и олигодендроглию. Этот слой нервной трубки является наиболее широким и насыщенным клеточными элементами.
Краевая вуаль наружный, самый светлый слой нервной трубки не содержит клеток, будучи выполненным их отростками, кровеносными сосудами и мезенхимой.
Особенностью клеток ганглиозной пластинки является то, что их дифференцировке предшествует период миграции в более или менее удаленные от первоначальной их локализации участки тела зародыша. Наиболее короткую миграцию претерпевают клетки, составляющие закладку спинномозговых узлов. Они спускаются на незначительное расстояние и располагаются по бокам от нервной трубки сначала в виде рыхлых, а затем более плотных крупных образований. У зародыша человека 6-8 нед развития спинномозговые узлы представляют собой очень большие образования, состоящие из крупных отростчатых нейронов, окруженных олигодендроглией. С течением времени нейроны спинномозговых ганглиев преобразуются из биполярных в псевдоуниполярные. Дифференцировка клеток внутри ганглиев происходит асинхронно.
Значительно более отделенную миграцию претерпевают те клетки, которые мигрируют из ганглиозной пластинки в ганглии пограничного симпатического ствола, ганглии превертебральной локализации, а также в мозговое вещество надпочечников. Особенно велика протяженность путей миграции нейробластов, вселяющихся в стенку кишечной трубки. Из ганглиозной пластинки они мигрируют по ветвям блуждающего нерва, достигают желудка, тонкой и наиболее краниальных частей толстой кишки, давая начало интрамуральным ганглиям. Именно столь длительным и сложным путем миграции структур, in situ управляющих процессом пищеварения, объясняется частота различного рода поражений этого процесса, возникающих как внутриутробно, так и после малейших нарушений диеты ребенка, особенно новорожденного или ребенка первых месяцев жизни.
Головной конец нервной трубки после ее замыкания очень быстро подразделяется на три расширения первичные мозговые пузыри. Сроки их образования, скорость клеточной дифференцировки и дальнейших преобразований у человека очень велики, Это позволяет рассматривать цефализацию опережающее и преимущественное развитие головного отдела нервной трубки как видовой признак человека.
Полости первичных мозговых пузырей сохраняются в мозге ребенка и взрослого в видоизмененной форме и образуют полости желудочков и сильвиева водопровода.
Самым ростральным отделом нервной трубки является передний мозг (prosencephalon); за ним следует средний (mesencephalon) и задний (rhombencephalon). В последующем развитии передний мозг делится на конечный (telencephalon), включающий полушария большого мозга и некоторые базальные ядра, и промежуточный (diencephalon). С каждой стороны промежуточного мозга вырастает глазной пузырь, формирующий нервные элементы глаза. Средний мозг сохраняется как единое целое, но в процессе развития в нем происходят значительные изменения, связанные с образованием специализированных рефлекторных центров, имеющих отношение к работе органов чувств: зрению, слуху, тактильной, болевой и температурной чувствительности.
Ромбовидный мозг делится на задний (metencephalon), включающий мозжечок и мост, и продолговатый (myelencephalon) мозг.
Одной из важных нейрогистологических характеристик развития нервной системы высших позвоночных является асинхронность дифференцировки ее отделов. Нейроны различных отделов нервной системы и даже нейроны в пределах одного центра дифференцируются асинхронно: а) дифференцировка нейронов вегетативной нервной системы значительно отстает от таковой в основных отделах соматической системы; б) дифференцировка симпатических нейронов несколько отстает от развития парасимпатических.
Раньше всего происходит созревание продолговатого и спинного мозга, позже морфологически и функционально развиваются ганглии ствола головного мозга, подкорковые узлы, мозжечок и кора больших полушарий. Каждое из этих образований проходит определенные этапы функционального и структурного развития. Так, в спинном мозге раньше созревают элементы в области шейного утолщения, а затем идет постепенное развитие клеточных структур в каудальном направлении; первыми дифференцируются спинальные мотонейроны, позже чувствительные нейроны и в последнюю очередь вставочные нейроны и проводящие межсегментные пути. Ядра стволовой части головного мозга, промежуточный мозг, подкорковые ганглии, мозжечок и отдельные слои коры большого мозга структурно развиваются также в определенной последовательности и в тесной связи друг с другом. Рассмотрим развитие отдельных областей нервной системы.
-
 «Комплексный подход к работе учителя – логопеда и педагога – психолога с детьми, имеющими речевые нарушения Характеристика изменений в
образовательном процессе
«Комплексный подход к работе учителя – логопеда и педагога – психолога с детьми, имеющими речевые нарушения Характеристика изменений в
образовательном процессе
-
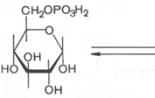 Бескислородное окисление глюкозы включает два этапа
Бескислородное окисление глюкозы включает два этапа
-
 Иисус навин кто он. Иисус сын навин. Вхождение в Землю Обетованную
Иисус навин кто он. Иисус сын навин. Вхождение в Землю Обетованную
-
 Что делать если заказ не пришел с eBay?
Что делать если заказ не пришел с eBay?









